SEP SNEST DGEST
UNIDAD VII:
TRANSCRIPCIÓN GENÉTICA
QUE PRESENTA:
Arisbeth Salgado Mendoza
09930054
Lic. BIOLOGIA VI "A"
Ciudad Altamirano, Gro. México. MAYO del 2012
INTRODUCCIÓN:

La traducción es el
segundo proceso de la síntesis proteica (parte del proceso general de la expresión génica).
La traducción ocurre tanto en el citoplasma, donde se encuentran los ribosomas, como también en el retículo endoplasmático rugoso (RER). Los ribosomas están formados
por una subunidad pequeña y una grande que rodean al ARNm. En la traducción, el ARN mensajero se
decodifica para producir un polipéptido específico de acuerdo con las reglas
especificadas por el código genético. Es
el proceso que convierte una secuencia de ARNm en una cadena de aminoácidos
para formar una proteína. Es necesario que la traducción venga precedida de un
proceso de transcripción. El proceso de traducción tiene cuatro fases:
activación, iniciación, elongación y terminación (entre todos describen el
crecimiento de la cadena de aminoácidos, o polipéptido, que es el producto de
la traducción).
En la activación, el aminoácido (AA) correcto se une al ARN de transferencia (ARNt) correcto. Aunque técnicamente esto no
es un paso de la traducción, es necesario para que se produzca la traducción.
El AA se une por su grupo carboxilo con el OH 3' del ARNt mediante un enlace de
tipo éster. Cuando el ARNt está enlazado con un aminoácido, se dice que está "cargado".
La iniciación supone que la subunidad pequeña del ribosoma se enlaza con el
extremo 5' del ARNm con la ayuda de factores de iniciación (FI), otras
proteínas que asisten el proceso. La elongación ocurre cuando el siguiente
aminoacil-ARNt (el ARNt cargado) de la secuencia se enlaza con el ribosoma
además de con un GTP y un factor de elongación. La terminación del polipéptido
sucede cuando la zona A del ribosoma se encuentra con un codón de parada (sin
sentido), que son el UAA, UAG o UGA. Cuando esto sucede, ningún ARNt puede
reconocerlo, pero el factor de liberación puede reconocer los codones sin
sentido y provoca la liberación de la cadena polipeptídica. La capacidad de
desactivar o inhibir la traducción de la biosíntesis de proteínas se utiliza en antibióticos como la anisomicina, la cicloheximida, el cloranfenicol y la tetraciclina.
El ARNm porta
información genética codificada
en forma de secuencia de ribonucleótidos desde los cromosomas hasta los
ribosomas. Los ribonucleótidos son "leídos" por la maquinaria
traductora en una secuencia de tripletes de nucleótidos llamados codones. Cada uno de estos tripletes codifica
un aminoácido específico. El ribosoma y las moléculas de ARNt
traducen este código para producir proteínas. El ribosoma es una estructura con varias subunidades que contiene ARNr y proteínas. Es la "fábrica" en la que se
montan los aminoácidos para formar proteínas. El ARNt son pequeñas cadenas de
ARN no codificador (de 74-93 nucleótidos) que transportan aminoácidos al
ribosoma. Los ARNt tienen un lugar para anclarse al aminoácido, y un lugar
llamado anticodón. El anticodón es un triplete de ARN complementario al
triplete de ARNm que codifica a su aminoácido. La aminoacil-ARNt sintetasa (una enzima) cataliza el enlace entre los ARNt específicos y los aminoácidos que concuerdan con sus
anticodones. El producto de esta reacción es una molécula de aminoacil-ARNt.
Esta aminoacil-ARNt viaja al interior del ribosoma, donde los codones de ARNm
se enfrentan con los anticodones específicos del ARNt mediante el emparejamiento de bases. Luego se utilizan
los aminoácidos que portan los ARNt para montar una proteína. La energía
requerida para traducir proteínas es significativa. Para una proteína que
contenga n aminoácidos, el número de enlaces
fosfato de alta energía necesarios para traducirla es 4n-1. Es también el proceso mediante el que los
ribosomas utilizan la secuencia de codones del ARNm para producir un
polipéptido con una secuencia particular de aminoácidos.
OBJETIVOS:
- Conocerá los eventos de síntesis proteica, la especificidad de la maquinaria enzimática y su implicación en la farmacología (inhibición de la síntesis proteica por antibióticos).
METODOLOGÍA:
En esta unidad el profesor nos fue presentando paso a paso como se daba el proceso de traducción del ARN mensajero nos mostró por medio de vídeos y fuimos analizando este proceso por el cual nosotros y vamos anotando y analizando cada paso para así aprender mas.
7.1 EL CÓDIGO GENÉTICO
Una vez que Crick (1958) propuso la
Hipótesis de la Secuencia ("existe una relación entre la ordenación lineal
de nucleótidos en el ADN y la ordenación lineal de aminoácidos en los
polipéptidos"), la comunidad científica la admitió y se plantearon dos
preguntas:
- ¿Existe
algún código o clave que permite pasar de la secuencia de nucleótidos en
el ADN a la secuencia de aminoácidos en las proteínas?
- ¿Cómo
se convierte la información contenida en la secuencia de ADN en una
estructura química de proteína?
La primera pregunta conlleva el estudio
del desciframiento del código genético y el estudio de sus características. La
segunda pregunta consiste en el estudio de los procesos genéticos de la
síntesis de proteínas: la transcripción y la traducción.
Las características del
código genético fueron establecidas experimentalmente por Fancis Crick, Sydney
Brenner y colaboradores en 1961. Las principales características del código
genético son las siguientes:


Sydney Brenner Francis crick
- El código está organizado en tripletes o codones: Cada tres nucleótidos
(triplete) determinan un aminoácido.
- El código genético es degenerado: Existen más tripletes
o codones que aminoácidos, de forma que un determinado aminoácido puede
estar codificado por más de un triplete.
- El código genético es no solapado o sin superposiciones: Un nucleótido
solamente pertenece a un único triplete.
- La lectura es "sin comas": El cuadro de lectura
de los tripletes se realiza de forma continua "sin comas" o sin
que existan espacios en blanco.
- El código genético nuclear es universal: El mismo triplete en
diferentes especies codifica para el mismo aminoácido. La principal
excepción a la universalidad es el código genético mitocondrial.
Si cada nucleótido determinara un
aminoácido, solamente podríamos codificar cuatro aminoácidos diferentes ya que
en el ADN solamente hay cuatro nucleótidos distintos. Cifra muy inferior a los
20 aminoácidos distintos que existen.
Si cada dos nucleótidos codificarán un
aminoácido, el número total de dinucleótidos distintos que podríamos conseguir
con los cuatro nucleótidos diferentes (A, G, T y C) serían variaciones con
repetición de cuatro elementos tomados de dos en dos VR4,2 = 42 =
16. Por tanto, tendríamos solamente 16 dinucleótidos diferentes, cifra inferior
al número de aminoácidos distintos que existen (20).
Si cada grupo de tres nucleótidos
determina un aminoácido. Teniendo en cuenta que existen cuatro nucleótidos
diferentes (A, G, T y C), el número de grupos de tres nucleótidos distintos que
se pueden obtener son variaciones con repetición de cuatro elementos (los
cuatro nucleótidos) tomados de tres en tres: VR4,3 = 43 =
64. Por consiguiente, existe un total de 64 tripletes diferentes, cifra más que
suficiente para codificar los 20 aminoácidos distintos.
Como hemos dicho anteriormente existen
64 tripletes distintos y 20 aminoácidos diferentes, de manera que un aminoácido
puede venir codificado por más de un codón. Este tipo de código se denomina
degenerado. Wittmann (1962) induciendo sustituciones de bases por desaminación
con nitritos, realizó sustituciones de C por U y de A por G en el ARN del virus
del mosaico del tabaco (TMV), demostrando que la serina y la isoleucina estaban
determinadas por más de un triplete.
Las moléculas encargadas de transportar
los aminoácidos hasta el ribosoma y de reconocer los codones del ARN mensajero
durante el proceso de traducción son los ARN transferentes (ARN-t). Los ARN-t
tienen una estructura en forma de hoja de trébol con varios sitios
funcionales:
- Extremo
3': lugar de unión al aminoácido (contiene siempre la secuencia ACC).
- Lazo
dihidrouracilo (DHU): lugar de unión a la aminoacil ARN-t sintetasa o
enzimas encargadas de unir una aminoácido a su correspondiente ARN-t.
- Lazo
de T ψ C: lugar de enlace al ribosoma.
- Lazo
del anticodón: lugar de reconocimiento de los codones del mensajero.
Normalmente el ARN-t adopta
una estructura de hoja de trébol plegada en forma de L o forma de boomerang.



Estructura ARN transferente
|
Estructura ARN transferente
|
Estructura ARN transferente
|
Los ARN-t suelen presentar bases
nitrogenadas poco frecuentes como son la pseudouridina (ψ), metilguanosina
(mG), dimetilguanosina (m2G), metilinosina (mI) y dihidrouridina
(DHU, UH2).
El que realiza el reconocimiento del
codón correspondiente del ARN-m es el anticodón del ARN-t y no el aminoácido.
Mediante un experimento se demostró que era posible transformar el
cisteinil-ARN-t mediante tratamiento con hidruro de níquel en alanil-ARN-t.
Este tratamiento convierte la cisteína en alanina. De esta manera se consiguió
un ARN-t específico de cisteina que en lugar de llevar unida cisteina llevaba
unida alanina. Cuando se empleo este ARN-t híbrido para sintetizar proteínas se
pudo comprobar que en el lugar en el que debía aparecer cisteina en la
secuencia del polipéptido aparecía alanina. Por tanto, el que llevaba a cabo el
reconocimiento del codón del ARN-m era el anticodón del ARN-t y no el
aminoácido.
La degeneración de l código se explica
teniendo en cuenta dos motivos:
- Algunos
aminoácidos pueden ser transportados por distintas especies moleculares
(tipos) de ARN transferentes (ARN-t) que contienen distintos anticodones.
- Algunas
especies moleculares de ARN-t pueden incorporar su aminoácido específico
en respuesta a varios codones, de manera que poseen un anticodón que es
capaz de emparejarse con varios codones diferentes. Este emparejamiento
permisivo se denomina Flexibilidad de la 3ª base del anticodón o
tambaleo.
Flexibilidad de la 3º base
del anticodón, tambaleo: La tercera base del anticodón, la que ocupa la posición 5' no está
especialmente confinada de forma que en algunos casos puede emparejarse con
distintas bases del codón. En la siguiente gráfica se observa que cuando en la
posición 5' del anticodón hay una G, esta guanina puede emparejarse con un
uracilo (U) de la posición 3' del codón o con una citosina (C).
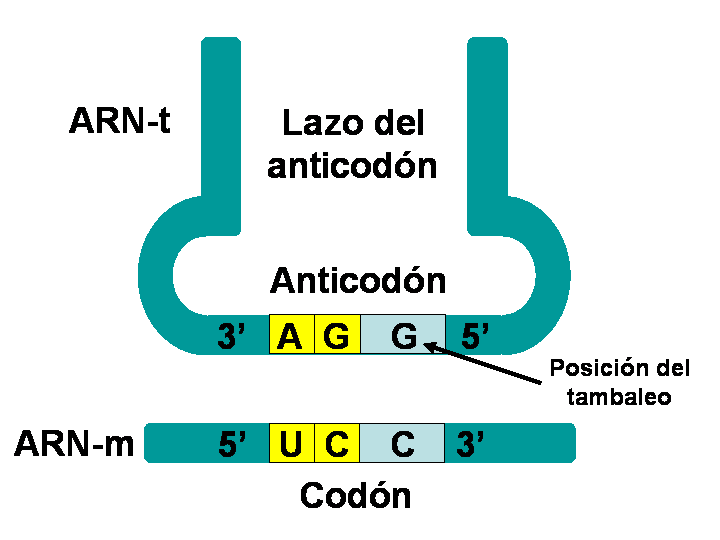

Flexibilidad
de la tercera base del anticodón, tambaleo.
EL CÓDIGO
GENÉTICO ES NO SOLAPADO O SIN SUPERPOSICIONES


Diferencias entre un código solapado y uno no
solapado
|
Código solapado: restricciones en la secuencia de
aminoácidos
|
Otra forma de comprobar que el código
es sin superposición es que no hay ninguna restricción en la secuencia de
aminoácidos de las proteínas, de manera, que un determinado aminoácido puede ir
precedido o seguido de cualquiera de los 20 aminoácidos que existen. Si dos
codones sucesivos compartieran dos nucleótidos, cualquier triplete solamente
podría ir precedido o seguido por cuatro codones determinados. Por
consiguiente, si el código fuera superpuesto, un aminoácido determinado
solamente podría ir precedido o seguido de otros cuatro aminoácidos como mucho.
Teniendo en cuenta que la lectura se
hace de tres en tres bases, a partir de un punto de inicio la lectura se lleva
a cabo sin interrupciones o espacios vacíos, es decir, la lectura es seguida
"sin comas". De manera, que si añadimos un nucleótido
(adición) a la secuencia, a partir de ese punto se altera el cuadro de lectura
y se modifican todos los aminoácidos. Lo mismo sucede si se pierde (deleción)
un nucleótido de la secuencia. A partir del nucleótido delecionado se altera el
cuadro de lectura y cambian todos los aminoácidos. Si la adición o la deleción
es de tres nucleótidos o múltiplo de tres, se añade un aminoácido o más de uno
a la secuencia que sigue siendo la misma a partir del la última adición o
deleción. Una adición y una deleción sucesivas vuelven a restaurar el cuadro de
lectura.
7.2 EL PAPEL DEL ARN EN LA SÍNTESIS DE PROTEÍNAS.
El
ARN mensajero es el que lleva la información para la síntesis de proteínas, es decir, determina el orden en que se unirán los
aminoácidos
La
síntesis de proteínas o traducción tiene
lugar en los ribosomas del
citoplasma celular. Los aminoácidos son transportados por el ARN de transferencia (ARNt) ,
específico para cada uno de ellos, y son llevados hasta el ARN mensajero (ARNm), dónde se aparean el codón de éste y elanticodón del ARN de
transferencia, por complementariedad de bases, y de ésta forma se sitúan
en la posición que les corresponde.
Una
vez finalizada la síntesis de una proteína, el ARN mensajero queda libre y
puede ser leído de nuevo. De hecho, es muy frecuente que antes
de que finalice una proteína ya está comenzando otra, con lo cual, una
misma molécula de ARN mensajero, está siendo utilizada por varios ribosomas simultáneamente.
·
Los
ARNt desempeñan un papel central en la síntesis de las proteínas
La
síntesis proteica tiene lugar en el ribosoma, que se arma en el citosol a partir de dos
subunidades riborrucleo proteicas provenientes del nucléolo. En el ribosoma el ARN mensajero (ARNm) se traduce
en una proteína, para lo cual se requiere también la intervención de los ARN de transferencia (ARNt). El trabajo de los ARNt consiste en tomar del citosol a
los aminoácidos y conducirlos al ribosoma en el orden marcado por los
nucleótidos del ARNm, que son los moldes del sistema
La
síntesis de las proteínas comienza con la unión entre sí de dos aminoácidos y
continúa por el agregado de nuevos aminoácidos -de a uno por vez- en uno
extremos de la cadena.
Como
se sabe la clave de la traducción reside en el código genético, compuesto por combinaciones de tres
nucleótidos consecutivos -o tripletes-
en el ARNm. Los distintos tripletes se relacionan específicamente con tipos de
aminoácidos usados en la síntesis de las proteínas.
Cada
triplete constituye un codón:
existen en total 64 codones, 61 de los cuales sirven para cifrar aminoácidos y
3 para marcar el cese de la traducción. Tal cantidad deriva de una relación matemática simple: los cuatro nucleótidos (A, U, C y
G)se combinan de a tres, por lo que pueden generarse 64 (43).
Dado
que existen más codones, (61) que tipos de aminoácidos (20), casi todos
pueden ser reconocidos por más de un codón, por lo que algunos tripletes a como
"sinónimos". Solamente el triptófano y la metionina -dos de los
aminoácidos menos frecuentes en las proteínas - son codificados, cada uno, por
un solo codón
Fig. A-1. Los dibujos ilustran cuatro de los seis codones que codifican
al aminoácido leucina (Leu). Los dos de la izquierda se aparean con un mismo
anticodón, igual que el par de codones de la derecha. Ello es posible
porque la tercera base de los codones suele ser "adaptable ", es
decir, puede establecer uniones con una base no complementaria.
 Generalmente
los codones que representan a un mismo aminoácido se parecen entre sí y es
frecuente que difieran sólo en el tercer nucleótido. La baja especificidad de
este nucleótido ha llevado a decir que existe una "degeneración" en tercera base de la
mayoría de los codones. Resta agregar que el número de codones en el ARNm
determina la longitud de la proteína
Generalmente
los codones que representan a un mismo aminoácido se parecen entre sí y es
frecuente que difieran sólo en el tercer nucleótido. La baja especificidad de
este nucleótido ha llevado a decir que existe una "degeneración" en tercera base de la
mayoría de los codones. Resta agregar que el número de codones en el ARNm
determina la longitud de la proteína
7.2.1 TIPOS DE ARN
El ARN mensajero (ARNm) es el tipo de ARN que lleva la
información del ADN a los ribosomas, el lugar de la síntesis de proteínas. La secuencia
de nucleótidos del ARNm determina la secuencia de aminoácidos de la proteína. Por
ello, el ARNm es denominado ARN codificante.
No obstante, muchos ARN no codifican proteínas, y reciben
el nombre de ARN no codificantes;
se originan a partir de genes propios (genes ARN), o son los intrones rechazados
durante el proceso de splicing.
Son ARN no codificantes el ARN de transferencia (ARNt) y el ARN ribosómico (ARNr), que son elementos
fundamentales en el proceso de traducción, y
diversos tipos de ARN reguladores.
Ciertos ARN no codificantes, denominados ribozimas, son capaces de catalizar reacciones químicas como cortar y unir
otras moléculas de ARN, o formar enlaces peptídicosentre aminoácidos en el ribosoma durante la síntesis de proteínas.
ARN implicados en la
síntesis de proteínas
§
ARN
mensajero. El ARN
mensajero (ARNm o RNAm) lleva la información sobre la secuencia de aminoácidos
de la proteína desde el ADN, lugar en que está inscrita, hasta el ribosoma, lugar
en que se sintetizan las proteínas de la célula. Es, por tanto, una molécula
intermediaria entre el ADN y la proteína y el apelativo de
"mensajero" es del todo descriptivo. En eucariotas, el ARNm se
sintetiza en el nucleoplasma del núcleo celular y de allí accede al citosol, donde se hallan
los ribosomas, a través de los poros de la envoltura nuclear.
§
ARN de transferencia. Los ARN de transferencia (ARNt o
tRNA) son cortos polímeros de unos 80 nucleótidos que transfiere un aminoácido
específico al polipéptido en crecimiento; se unen a lugares
específicos del ribosoma durante la traducción. Tienen un sitio específico para
la fijación del aminoácido (extremo 3') y un anticodón formado por un triplete de nucleótidos
que se une al codón complementario del
ARNm mediante puentes de hidrógeno.
§
ARN
ribosómico. El ARN
ribosómico (ARNr o RNAr) se halla combinado con proteínas para formar los
ribosomas, donde representa unas 2/3 partes de los mismos. En procariotas, la
subunidad mayor del ribosoma contiene dos moléculas de ARNr y la subunidad
menor, una. En los eucariotas, la subunidad mayor contiene tres moléculas de
ARNr y la menor, una. En ambos casos, sobre el armazón constituido por los ARNr
se asocian proteínas específicas. El ARNr es muy abundante y representa el 80%
del ARN hallado en el citoplasma de las células eucariotas. Los ARN ribosómicos son el componente
catalítico de los ribosomas; se encargan de crear los enlaces peptídicos entre
los aminoácidos del polipéptido en formación durante la síntesis de proteínas;
actúan, pues, como ribozimas.
ARN
reguladores
Muchos
tipos de ARN regulan la expresión
génica gracias a que son complementarios de regiones específicas del ARNm o de
genes del ADN. .
§
ARN de interferencia. Los ARN interferentes (ARNi o iRNA)
son moléculas de ARN que suprimen la expresión de genes específicos mediante
mecanismos conocidos globalmente como ribointerferencia o interferencia por ARN. Los ARN
interferentes son moléculas pequeñas (de 20 a 25 nucléotidos) que se generan
por fragmentación de precursores más largos. Se pueden clasificar en tres
grandes grupos:
§
Micro ARN. Los micro ARN (miARN o RNAmi) son
cadenas cortas de 21 ó 22 nucleótidos hallados en células eucariotas que se
generan a partir de precursores específicos codificados en el genoma. Al transcribirse, se pliegan en horquillas
intramoleculares y luego se unen a enzimas formando un complejo efector que
puede bloquear la traducción del ARNm o acelerar su degradación comenzando por
la eliminación enzimática de la cola poli A.
§
ARN interferente pequeño. Los ARN interferentes pequeño (ARNip
o siARN), formados por 20-25 nucleótidos, se
producen con frecuencia por rotura de ARN virales, pero pueden ser también de origen endógeno. Tras la transcripción se ensambla en
un complejo proteico denominado RISC (RNA-induced silencing complex) que
identifica el ARNm complementario que es cortado en dos mitades que son
degradadas por la maquinaria celular, bloquean así la expresión del gen.
§
ARN
asociados a Piwi. Los ARN asociados a Piwi son cadenas
de 29-30 nucleótidos, propias de animales; se generan a partir de precursores
largos monocatenarios, en un proceso que es independiente de Drosha y Dicer. Estos ARN pequeños se asocian con una
subfamilia de las proteínas "Argonauta" denominada proteínas Piwi.
Son activos las células de la línea germinal; se cree que son un sistema defensivo
contra los transposones y que juegan algún papel en la gametogénesis.
§
ARN
antisentido. Un ARN
antisentido es la hebra complementaria (no codificadora) de un hebra ARNm
(codificadora). La mayoría inhiben genes, pero unos pocos activan la
transcripción. El ARN antisentido
se aparea con su ARNm complementario formando una molécula de doble hebra que
no puede traducirse y es degradada enzimáticamente. La introducción de un transgen codificante para un ARNm antisentido
es una técnica usada para bloquear la expresión de un gen de interés. Un mARN
antisentido marcado radioactivamente puede usarse para mostrar el nivel de
transcripción de genes en varios tipos de células. Algunos tipos estructurales
antisentidos son experimentales, ya que se usan como terapia antisentido.
§
ARN largo no codificante. Muchos ARN largos no codificantes
(ARNnc largo o long ncARN) regulan la expresión génica en eucariotas; uno de ellos es el Xist que recubre uno de los dos cromosomas X en las hembras de los mamíferos inactivándolo (corpúsculo
de Barr).
§
Riboswitch. Un riboswitch es una parte del ARNm (ácido ribonucleico
mensajero) al cual pueden unirse pequeñas moléculas que afectan la actividad
del gen. Por tanto, un ARNm que
contenga un riboswitch está directamente implicado en la regulación de su
propia actividad que depende de la presencia o ausencia de la molécula
señalizadora. Tales riboswitchs se hallan en la región no traducida 5'
(5'-UTR), situada antes del codón de inicio (AUG), y/o en la región no
traducida 3' (3'-UTR), también llamada secuancia de arrastre, situada entre el codón de terminación
(UAG, UAA o UGA) y la cola poli A.
7.2.2 ESTRUCTURA RIBOSOMAL

Los
ribosomas son orgánulos sin membrana, sólo visibles al microscopio electrónico
debido a su reducido tamaño (29 nm en células procariotas y 32 nm en
eucariotas). Están en todas las células vivas. Su función es ensamblar
proteínas a partir de la información genética que le llega del ADN transcrita
en forma de ARN mensajero (ARNm).
ESTRUCTURA*
- · El ribosoma consta de dos partes, la subunidad mayor y una menor, estas salen del núcleo por separado.
- · El ribosoma procarionta tiene un coeficiente de sedimentación de 70s y está formado por dos subunidades de 50s y 30s. Se puede encontrar en el citoplasma donde recibe el nombre de polisoma o polirribosa.
- · El ribosoma eucariota tiene un coeficiente de sedimentación de 80s, uno de 60s y otra de 40s Este se puede encontrar unido al retículo endoplasmático rugoso.
- · La unión de ambas subunidades se realiza en presencia de una concentración 0.001 M de iones Mg, si esta concentración disminuye se produciría la separación de las dos subunidades, es por lo tanto un proceso reversible.
- · Si la concentración molar de los iones Mg aumenta hasta 10 veces, se produce la unión de dos ribosomas para dar lugar a los dímeros.
- · La ultraestructura del ribosoma es muy compleja, está formado por ARN ribosómico y proteinas.
- · Los ribosomas son responsables del aspecto granuloso del citoplasma de las células. Es el orgánulo más abundante, hay varios millones por célula.
BIOGÉNESIS
DE LOS RIBOSOMAS
Dada
la gran complejidad que tienen los ribosomas cabe pensar que hay un número
bastante elevado de genes que participan en su formación. La biogénesis varía
según sea eucariotas o procariotas.
En
procariotas los estudios se realizaron sobre E.Coli, a la cual se la hacía
crecer en un medio rico en uracilo rico en carbono 14. Se comprobó que el
material que formaba parte de los ribosomas aparecen dos tipos de partículas de
coeficientes 8s y 20s. Al poco tiempo desaparecían y aparecían otras de 26s y
40s, estudiándolas se vio que de 26s tenían un ARN de 16s y 20s
respectivamente. Al poco tiempo desaparecían y aparecían las subunidades mayor
y menor de los ribosomas de células procariotas.
En
eucariotas su origen está relacionado con los nucleolos. Un precursor sería el
ARN ribosómico 45s, al poco tiempo aparecían dos ARN marcados uno de 32s y otro
de 18s. El 18s pasaba rápidamente al citoplasma, el 32s permanecía cierto
tiempo en el interior hasta que desaparecía
FUNCION*
La
función de los ribosomas es la síntesis de proteínas. Este es el proceso
mediante el cual el mensaje contenido en el ADN nuclear, que ha sido
previamente transcrito en un ARN mensajero, es traducido en el citoplasma,
juntamente con los ribosomas y los ARN de transferencia que transportan a los
aminoácidos, para formar las proteínas celulares y de secreción.
El
ribosoma lee el ARN mensajero y ensambla la proteína con los aminoácidos
suministrados por los ARN de transferencia, este proceso se denomina síntesis de
proteínas.
Los
polisomas se encargan de sintetizar proteínas de localización celular, mientras
que los ribosomas del RER se encargan de sintetizar proteínas de exportación, o
sea que se irán de la célula hacia otro lugar donde se necesite.
Para
la síntesis de proteínas se requiere la unión de una subunidad mayor y otra
menor. Al terminar de fabricar las proteínas, se separan. Cuando vuelven a
sintetizar se unen dos diferentes.
Para
la síntesis de proteínas los ribosomas se asocian en grupos mediante un
filamento de unos 2 nm. de espesor. Estos ribosomas asociados se denominan
polisomas, y suelen adoptar una figura en espiral. La subunidad menor queda
hacia el interior de la espiral.
Los
ribosomas forman polisomas para realizar cualquier tipo de síntesis proteicas:
tanto la efectuada por los ribosomas libres como la realizada por los asociados
a membranas (RER). En el RER la subunidad mayor es la que se adosa a la
membrana.
El
número de ribosomas que forman un polisoma y la longitud de ARN-m que los une
varía según el peso molecular de las proteínas que se va a sintetizar.
Para
la síntesis de proteínas, los ribosomas recorren el ARNm desde un extremo a
otro. Por cada tres nucleótidos recorridos, incorporan un aminoácido a la cadena
de proteínas que están sintetizando; aminoácidos que les proporciona el ARNt .
Cuando
han completado el recorrido, los ribosomas se liberan del ARNm y suelta la
proteína ya terminada. Mientras se esté sintetizando proteínas, por cada
ribosoma que abandona el polisoma en el extremo final, otro se incorpora en el
inicial, de modo que el ritmo de trabajo del polisoma siempre es complejo.
El
ARNt y la cadena de aminoácidos que se está formando se encuentran en un canal
situado en la subunidad mayor.
7.2.3 PROCARIOTICO

En bacterias, la subunidad
50S es aproximadamente dos veces la masa de la subunidad 30S, y en ambas,
aproximadamente 2/3 de su peso se debe al ARNr mientras que el resto del peso es
atribuible a la parte proteica del ribosoma.
La subunidad 30S tiene una
forma más o menos trapezoidal cuando se tiene una vista frontal desde la zona
de interacción de subunidades (Figura 1.2) y es uniformemente estrecha cuando se
tiene una visión lateral. En la subunidad 30S se distinguen la cabeza y cuerpo.
Del cuerpo salen dos lóbulos hacia arriba, denominados “plataforma” y “espalda”.
A la hendidura que se forma entre la
plataforma y la cabeza se la conoce por ser el centro de decodificación.
La subunidad 50S tiene una
forma aproximadamente hemisférica enfrentada a la subunidad 30S por su lado
liso. Desde la región de interacción entre subunidades se distinguen 3 protuberancias:
el tallo L7/L12 que se extiende a lo largo de su base, la protuberancia central
y el tallo L1. El tallo L7/L12, formado por las proteínas diméricas L7/L12, a
veces puede estar ausente en mapas de microscopía electrónica debido a su
elevada flexibilidad. La protuberancia central está formada, en su mayor parte,
por el ARNr 5S. Al contrario que el tallo L7/L12, el tallo L1 está siempre bien definido,
formado por la proteína ribosomal L1 y parte del ARNr En un ribosoma 70S, las
dos subunidades se enfrentan de tal manera que se crea un espacio de morfología
complejo (Figura 1.3): el espacio entre subunidades, por donde irán circulando
los
ARNt durante la traducción.
El ensamblaje se mantiene mediante diversos puentes que se crean entre
subunidades. Existe un total de 12 puentes entre subunidades creados por más de
30 interacciones individuales y la mayoría de ellos involucran contactos entre ARNr (Yusupov et al., 2001).
Sin embargo, los puentes que se crean entre subunidades no confieren al
ribosoma 70S una estructura totalmente rígida. La naturaleza dinámica del
proceso de traducción implica que el ribosoma sea una estructura flexible con
componentes móviles que posibiliten su función (Spirin, 1969).
7.2.4 EUCARIOTICO

Los ribosomas de eucariotas
tienen aproximadamente 80S y un peso molecular de unos 4.200 kDa, aunque
existen pequeñas diferencias entre especies. En eucariotas el rRNA, que es el 70%
de todo el RNA de la célula, constituye el 40% del peso del ribosoma. La
composición en bases de este rRNA es diferente a la del rRNA de procariotas. La
subunidad mayor (60S) tiene un RNA de 28S, otro de 5,8S y otro de 5S. La
subunidad menor tiene un RNA de 18S, aunque hay pequeñas variaciones
dependiendo del organismo en particular. Las proteínas ribosomales difieren de
las de procariotas. Hay 33 proteínas en la subunidad menor y 49 en la subunidad
mayor, y la mayoría de las proteínas de cada subunidad (o posiblemente
todas) son diferentes de las presentes
en la otra subunidad. La síntesis proteica de estos ribosomas es inhibida por agentes
que también inhiben la síntesis proteica en procariotas (como puromicina y
actinomicina D) y por inhibidores específicos, como la alfa-amanitina, la anisomicina
y la cicloheximida.
En eucariotas también hay
ribosomas en las mitocondrias y en los plastos que se asemejan a los de
procariotas y, como ellos, son inhibidos por cloranfenicol. Los ribosomas de
los plastos son muy semejantes a los ribosomas procarióticos y, como éstos,
tienen 70S. Los ribosomas mitocondriales difieren de ellos en ciertos aspectos.
En células animales, los ribosomas mitocondriales son menores que los ribosomas
de bacterias y varían algo de unas especies a otras (entre 55S y 60S). Por el
contrario, en levaduras y ciliados, los ribosomas mitocondriales suelen ser mayores
que los de procariotas. En la subunidad mayor presentan un RNA de 4S,
equivalente al de 5S presente en los ribosomas de procariotas y en los
ribosomas citoplasmáticos de eucariotas.
En E. coli y otros
procariotas, aun cuando el extremo 3' de una cadena de mRNA está siendo
transcripto, se están uniendo ribosomas cerca de su extremo 5'. En el lugar
donde la cadena de mRNA está en contacto con un ribosoma, se unen tRNAs
temporalmente a la cadena de mRNA. Esta unión ocurre por apareamiento de bases
complementarias entre el codón de mRNA y el anticodón de tRNA. Cada molécula de
tRNA lleva el aminoácido específico requerido por el codón de mRNA, al cual se
une el tRNA. Así, siguiendo la secuencia dictada originalmente por el DNA, las
unidades de aminoácidos son alineadas una tras otra y, a medida que se forman
los enlaces peptídicos entre ellas, se unen en una cadena polipeptídica.
Esquema general del flujo de
información en procariotas y eucariotas: a) En procariotas, el RNA se
transcribe a partir de una molécula de DNA circular y, a medida que ocurre la
transcripción, se produce la traducción en el mismo compartimiento. b) En
eucariotas, la transcripción ocurre en el núcleo y el RNA, luego de sufrir un
procesamiento, se dirige al citoplasma donde se produce la síntesis de
proteínas. Como se vio en el capítulo 5, algunas proteínas son sintetizadas en
los ribosomas libres y otras en los que están adheridos al retículo
endoplásmico.

Tres etapas en la síntesis
de proteínas en procariotas

La síntesis de proteínas
ocurre en varias etapas: a) Iniciación. La subunidad ribosómica más pequeña se
une al extremo 5' de una molécula de mRNA. La primera molécula de tRNA, que
lleva el aminoácido modificado fMet, se acopla con el codón iniciador AUG de la
molécula de mRNA. La subunidad ribosómica más grande se ubica en su lugar, el
complejo tRNA-fMet ocupa el sitio P (peptídico). El sitio A (aminoacil) está
vacante. El complejo de iniciación está completo ahora.

Un segundo tRNA, con su
aminoácido unido, se coloca en el sitio A y su anticodón se acopla con el mRNA.
Se forma un enlace peptídico entre los dos aminoácidos reunidos en el ribosoma.
Al mismo tiempo, se rompe el enlace entre el primer aminoácido y su tRNA. El
ribosoma se mueve a lo largo de la cadena de mRNA en una dirección 5' a 3', y
el segundo tRNA, con el dipéptido unido, se mueve desde el sitio A al sitio P,
a medida que el primer tRNA se desprende del ribosoma. Un tercer aminoacil-tRNA
se coloca en el sitio A y se forma otro enlace peptídico. La cadena peptídica
naciente siempre está unida al tRNA que se está moviendo del sitio A al sitio P
y el tRNA entrante que lleva el siguiente aminoácido siempre ocupa el sitio A.
Este paso se repite una y otra vez hasta que se completa el polipéptido.

Cuando el ribosoma alcanza
un codón de terminación (en este ejemplo UGA), el polipéptido se escinde del
último tRNA y el tRNA se desprende del sitio P. El sitio A es ocupado por un
factor de liberación que produce la disociación de las dos subunidades del
ribosoma.

A partir del DNA cromosómico
se transcriben: diferentes moléculas de rRNA que, combinadas con proteínas
específicas, forman los ribosomas; los diferentes tipos de moléculas de tRNA
correspondientes a los distintos aminoácidos y los mRNA, que llevan la
información para la secuencia de aminoácidos de las proteínas. Cuando un mRNA
se une a la subunidad menor del ribosoma, comienza el proceso de síntesis de
proteínas.
7.4 ETAPAS DE LA SÍNTESIS DE PROTEÍNAS EN ORGANISMOS EUCARIOTICOS.

Transcripción
Traducción
Iniciación
Elongación
o alargamiento
Terminación

El
ARN o ácido ribonucleico complementa las funciones del ADN, ya que interpreta
el mensaje hereditario y lo traduce en la formación de las proteínas que el
organismo requiere.
Cada célula de un organismo elabora sus proteínas de
acuerdo con la información que el ADN proporciona. No toda la información es
leída al mismo tiempo. Esto se puede compara con lo que ocurre
con una gran enciclopedia: sólo se abre en determinados capítulos y se copia de
acuerdo con las necesidades. Lo mismo ocurre con la información contenida en el
ADN y su utilización por la célula.
Al igual que el ADN, el ARN está formado por nucleótido,
pero existen diferencias esenciales entre ambos tipos de moléculas.
El ARN más abundante en las células es el ribosomal
(ARNr), porque forma los ribosomas, organelos donde se sintetizan las
proteínas; se encuentra en un porcentaje de 85 a 90%. El ARN de transferencia
(ARNt) representa del 5 al 7% del ARN total, mientras que el ARN mensajero
(ARNm) representa del 3 al 5%.
Las funciones de interpretación del mensaje genético por el ARN tienen
lugar en dos etapas:
Transcripción, durante la cual el ARN mensajero produce
una copia de un segmento de ADN, correspondiente a un gen, y esto conduce a la
elaboración de una proteína determinada.
Traducción, proceso que
se lleva a cabo en los ribosomas, cuando se incorporan los aminoácidos en el
orden preciso, de acuerdo con las "instrucciones" o información del
ARN mensajero
Transcripción
Esta se lleva a cabo con la intervención de la enzima ARN
polimerasa; la cual tiene la función de colocar nucleótidos de ARN en el orden
que corresponde al segmento de ADN que se está copiando.
El proceso muestra diferencias
según se realice en células procariontes o eucariontes. En las procariontes
existen tres etapas: iniciación, elongamiento o alargamiento y terminación o
finalización. Mientras que en las eucariontes, además de las tres anteriores,
se agrega una de maduración, en la que el ARN sufre modificaciones, como la
eliminación de las secuencias correspondientes a intrones, las cuales no son
necesarias en la etapa de síntesis de proteínas. Otra diferencia es que en
procariontes el proceso de transcripción se lleva a cabo en el citoplasma, y en
los eucariontes, dentro del núcleo.
Enseguida se explica cada una de las etapas:
Iniciación. La ARN polimerasa se une a una región
determinada del ADN llamada promotor.
Elongación o alargamiento. En esta etapa, se abre la
molécula de ADN en la zona donde se ha instalado la ARN polimerasa y empiezan a
colocarse los nucleótidos complementarios a la secuencia del ADN. La formación
del ARN va ocurriendo en dirección 5
3. Después que se han colocado unos treinta o cuarenta nucleótidos, se añade
una base modificada en la zona del inicio del ARNm, de la cual se dice que es
la "capucha" del ARNm, la cual le permitirá unirse más tarde al
ribosoma.
Finalización o terminación. Cuando la enzima ARN
polimerasa llega al mensaje de terminación de cadena, que es una determinada
secuencia en el mensaje del ADN, concluye la formación de la molécula de ARN
mensajero, y se agrega un segmento formado por doscientas unidades de adenina.
Este segmento se conoce como cola de poli-A, y ayudará a prolongar la vida
activa de la molécula de ARN mensajero.
Maduración. Se ha una molécula de pre-ARN mensajero. Lo
único que falta es eliminar las secuencias sin sentido, los intrones, para que
este quede totalmente terminado. Una enzima llamada ribozima se encarga de
cortar las secuencias de intrones y otra enzima pega y empalma los segmentos
que han quedado, lo cual da lugar finalmente al ARN maduro, que ya puede llevar
a cabo la síntesis de proteínas
Traducción
La síntesis de proteínas se conoce también como traducción, dado que es la transferencia de información
de los nucleótidos al de los aminoácidos, por lo que es considerada la
actividad sintética más compleja que ocurre en la célula. En tanto que otras
macromoléculas de las células se elaboran por medio de reacciones enzimáticas
relativamente directas, el ensamblado de una proteína requiere todos los
diferentes RNAt con sus aminoácidos unidos, ribosomas, RNA mensajeros y algunas
proteínas con varias funciones, cationes y GTP
La complejidad no es sorprendente si consideramos que la
síntesis de proteínas requiere la incorporación de cada uno de los diferentes
20 aminoácidos en la secuencia precisa dictada por un mensaje codificado
escrito en un lenguaje que
emplea distintos símbolos.
El proceso de traducción en las células eucariotas es
similar en las células procariotas, la diferencia principal es que la
traducción en células eucariotas implica numerosos factores proteínicos
solubles (no ribosómicos)
En este proceso de traducción intervienen los tres tipos
de ARN: ribosomal, de transferencia y mensajero.
ARN ribosomal. Los ribosomas están constituidos por ARN
ribosomal, y es en ellos donde se lleva a cabo el proceso de traducción, la
síntesis de proteínas. Cada ribosoma está formado por dos subunidades, una
menor que otra, que se ensamblan en el momento de inicio de la síntesis de
proteínas.
ARN de transferencia. Tiene forma de trébol, y en algunos
segmentos presenta una doble cadena. En una región de esta molécula se encuentra
el anticodón, secuencia de tres bases complementarias con las del ARNm. En otra
parte de la molécula, el ARN posee el espacio para unirse de manera específica
con un aminoácido determinado. Existen por lo menos cincuenta ARNt diferentes,
y cada uno de ellos posee un anticodón que se corresponde con un aminoácido
determinado. Durante la síntesis de proteínas, los ARNt son acarreadores de
aminoácidos.
ARN mensajero. Contiene la información que ha transcrito
del ADN la cual se lee por tripletes, por secuencias de tres letras; a cada una
de las cuales se le llama codón. Para cada codón hay un ARNt, con su respectivo
anticodón, de modo que en la síntesis de proteínas cada aminoácido queda
colocado de manera específica, de acuerdo con el mensaje genético del ARNm En
los eucariontes, la síntesis de proteínas se lleva a cabo en tres etapas
distintas: iniciación, elongación o alargamiento y terminación.
Iniciación
En eucariontes, la iniciación comienza cuando la
subunidad ribosómica pequeña reconoce primero el extremo 5´ del mensaje que
precede al casquete de metilguanosina, y luego se desplaza a lo largo del RNAm
hasta alcanzar una secuencia de nucleótidos (típicamente 5´-CCACCAUG-3´) que
contiene el tripleto AUG en un contexto en el cual se le puede reconocer como
codón inicial
A continuación, el primer ARNt iniciador se coloca en su
lugar y se aparea con el codón iniciador del ARNm. Este codón iniciador que
habitualmente es (5´)-AUG-(3´), se aparea en forma antiparalela con el
anticodón del ARNt (3´)-UAG-(5´). El ARNt iniciador entrante, que se une al
codón AUG, lleva una forma modificada del aminoácido metionina,
N-formilmetionina o fMet. Esta fMet será el primer aminoácido de la cadena
polipeptídica recién sintetizada que es rápidamente removido.
El ARNt iniciador está ubicado en el sitio P de la
subunidad mayor, uno de los dos sitios de unión para las moléculas de ARNt.
Luego, se liberan estos factores de iniciación y la subunidad ribosómica mayor
se une a la subunidad menor. La energía para este paso la suministra la
hidrólisis del trifosfato de guanosina
Elongación
o alargamiento
Una vez que el ribosoma completo se ha ensamblado en el
codón de inciación, comienza la etapa de elongación o alargamiento. Durante
esta etapa, elsitio A de un ribosoma será ocupado transitoriamente por
sucesivos aminoacil.ARNt.Los aminoacil-ARNt que ocupen el sitio A serán
aquellos cuyo anticodón sea complementario al codón que queda expuesto en ese
sitio. La entrada del aminoacil-ARNt al sitio A del ribosoma requiere su unión
previa con una proteína llamada factor de elongación, que en su forma activa
será unida al GTP. Al aparearse el ARNt con el ARNm, se dispara la hidrólisis
del GTP por parte del factor de elongación que luego se disocia, permitiendo
que el aminoacil-ARNt permanezca unido por un corto período al ARNm.
Cuando los sitios A como P están ocupados, una enzima, la
peptidil transferasa, que es parte de la subunidad mayor del ribosoma, cataliza
la formación de un enlace peptídico entre los dos aminoácidos, acoplando el
primero (fMet) al segundo. El primer ARNt, entonces se libera. El ribosoma se
mueve un codón a lo largo de la cadena de ARNm; en consecuencia, el segundo
ARNt, al cual ahora se encuentran acoplados la fMet y segundo aminoácido, se
transfiere de la posición A a la posición P. Un tercer aminoacil-ARNt se ubica
en la ahora vacante posición A, apareado al tercer codón del ARnm, y se repite
el paso. La posición P acepta al ARNt que carga con la cadena polipeptídica
creciente; la posición A acepta al ARNt que porta el nuevo aminoácido que será
añadido a la cadena.
A medida que el ribosoma se mueve a lo largo de la cadena
del ARNm, la porción iniciadora de la molécula de ARNm es liberada y otro
ribosoma puede formar con ella un complejo de iniciación. Un grupo de
ribosomas que leen la misma molécula de ARNm se conoce como polisoma
Terminación
Cuando el mensaje llega a un codón de terminación, ya no
hay ningún ARNt que tenga su anticodón, de manera que ya no se agrega otro
aminoácido a la cadena. Se libera el polipéptido formado, la proteína. Se
separan las dos subunidades del ribosoma y el ARNm es liberado
7.4.1 MODIFICACIÓN DE PROTEÍNAS POSTRADUCCION
- Para asumir actividad biologica, muchos polipeptidos nacientes tienen que ser modicados de forma covalente antes o despues del proceso de plegado.
- Las modicaciones posteriores a la traduccion tienen un gran impacto en la funcion de la proteina, al alterar el tamaño, hidrofobicidad y sobre todo la conformacion de la proteina.
- Las modicaciones pueden inuenciar directamente las interacciones proteina-proteina y la distribucion de proteinas a diferentes localidades subcelulares.
- El motivo [ST]-x-[RK] puede ser encontrado varias veces en casi todas las secuencias de proteinas. La mayoria de las predicciones basadas en este motivo de secuencia solo pueden estar equivocados produciendo tasas muy elevadas de falsos positivos.
- Para minimizar los resultados falsos positivos, se puede utilizar un proceso de aprendizaje estadistico llamado maquina de soporte vectorial (SVM) para aumentar la especificad de la predicción.
- Este es un metodo de clasicacion de datos similar al analisis discriminatorio lineal o cuadratico.
- Después de entrenar el algoritmo con suficientes características estructurales, esta habilitado para reconocer correctamente muchos patrones de modificaciones posteriores a la traduccion.
- AutoMotif, es un servidor web de predicción de motivos en secuencias de proteínas usando el enfoque de SVM. En este proceso la secuencia de consulta es dividida entre el número de fragmentos solapados, los cuales son alimentados en diferentes nucleos (similar a los nodos).
- Un hiperplano, el cual ha sido entrenado para reconocer motivos de secuencias de proteinas conocidas, separa los nucleos en diferentes clases.
- Cada separación es comparada con clases de motivos conocidas, de las cuales la mayor a estan relacionadas a modificaciones posteriores a la traducción.
- La mejor coincidencia con una clase conocida define el motivo funcional.
TAREA: ¡EXPLIQUE! ES POSIBLE QUE LA MAQUINA EUCARIOTES DE TRADUCCIÓN (RIBOSOMAS,FACTORES,ENZIMAS,RNAt) PUEDAN TRADUCIR EL RNAm DE UNA BACTERIA. ¡EXPLIQUE! IDENTIFICANDO LOS ION INCONVENIENTES QUE ENCONTRARAS.
nop..... ya que la traducción en ambas son similares pero encontramos varias diferencias que hacen que no pueda ser posible la realización.
- La primera seria por que los procariontes se da este proceso por fragmentos de proteínas. y las eucariotes no en forma circular.
- El segundo desacuerdo con esto es que en procariontes encontramos 3 sitios donde se puedes ser la traducción y en eucarionte solo dos sitios.
- El tercero seria que en procariontes es mas pequeño 50s. y los eucariotes es mas grande 80s.
- El cuarto seria que la metionina en procariontes debe ser formilada para empezar el proceso y en comparación de la eucariotes no es necesario que esta metionina se encuentre así que no es posible este proceso....
En esta unidad vimos como se realiza la traduccion en diferentes organimos (celula eucariota y procariota) a su vez las modificaciones de proteínas POSTRADUCCION por el código genético y sobre todos los diferentes tipos de ARN. poco a poco los fuimos analizando.
BIBLIOGRAFIA:
http://www.ucm.es/info/genetica/grupod/Codigo/Codigo%20genetico.htm
http://www.monografias.com/trabajos/sinteproteinas/sinteproteinas.shtml
http://www.mitecnologico.com/ibq/Main/EstructuraYFuncionDeLosRibosomas
http://www.argitalpenak.ehu.es/p291content/eu/contenidos/informacion/se_indice_tescvpdf/eu_tescvpdf/adjuntos/PATRICIA%20JULIAN%20TESIS.pdf
http://retina.umh.es/docencia/confsvivos/temas/Tema_11_Ribosomas.pdf
http://www.angelfire.com/magic2/bioquimica/sintesis_de_prote_nas.htm
http://www.monografias.com/trabajos68/sintesis-proteinas-eucariontes/sintesis-proteinas-eucariontes2.shtml
http://www.tamps.cinvestav.mx/~ertello/bioinfo/sesion18.pdf
No hay comentarios:
Publicar un comentario